- BIODÉMOGRAPHIE - Dynamique des populations
- BIODÉMOGRAPHIE - Dynamique des populationsLes caractères généraux des populations animales monospécifiques, envisagés principalement d’un point de vue quantitatif (répartition des classes d’âge, proportion des sexes, taux de croissance, de fécondité, de mortalité, etc.), ne sont pas fixés une fois pour toutes, mais ils sont susceptibles de variations plus ou moins importantes dans l’espace et le temps. La dynamique des populations a pour objet de décrire et de mesurer ces variations, puis d’en chercher les causes. Dans le cas où celles-ci échappent au contrôle de l’homme, le but est d’arriver à en prévoir les effets. Si en revanche elles peuvent être contrôlées par l’homme, le but sera d’indiquer les moyens de diriger l’évolution des populations dans un sens déterminé.HistoriqueLes variations d’abondance de certaines populations animales (rats, sauterelles, moustiques) ont frappé l’imagination de l’homme dès la plus haute antiquité. Ce n’est toutefois qu’à la fin du XIXe et au début du XXe siècle que des observations précises sur des essais de contrôle de la cochenille de l’oranger par une coccinelle, la destruction du pigeon migrateur américain, la disparition quasi totale du bison, la mortalité des poissons au large des côtes des Bermudes à la suite d’un hiver exceptionnellement rigoureux, la pullulation de lépidoptères forestiers ravageurs introduits aux États-Unis attirent l’attention des biologistes sur l’importance pratique et les incidences économiques des études quantitatives de populations.À partir de 1930, des expérimentations en laboratoire (G. F. Gause, 1935) et des analyses mathématiques (A. J. Lotka, 1934, V. Volterra, 1935, V. A. Kostitzin, 1934) viennent s’ajouter aux observations faites en milieu naturel. De nombreuses théories sont alors proposées, surtout par des entomologistes. Les unes insistent sur le rôle primordial des facteurs biotiques, surtout prédateurs et parasites (A. J. Nicholson, de 1954 à 1958, W. Edwards, de 1962 à 1970, J. J. Christian et D. E. Davis, 1964). D’autres considèrent comme plus importants les facteurs abiotiques, notamment climatiques (H. G. Andrewartha et L. C. Birch, 1954, W. R. Thompson, de 1929 à 1956). D’autres encore cherchent à concilier les deux tendances précédentes (A. Milne, 1957), font appel à des mécanismes génétiques (D. Chitty, de 1960 à 1970) ou introduisent des concepts cybernétiques (H. Wilbert, de 1962 à 1971).Depuis la Seconde Guerre mondiale, les biologistes des pêches (M. Graham, 1956, R. J. H. Beverton et S. J. Holt, 1957, W. E. Ricker, 1958, J. A. Gulland, de 1962 à 1971) ont donné une vigoureuse impulsion à l’étude des stocks exploités, particulièrement ceux des poissons. Les divers aspects théoriques ou appliqués de la dynamique des populations se développent d’autant plus rapidement que la vulgarisation des ordinateurs offre, pour traiter les données et tester les modèles, des facilités que les plus optimistes n’auraient pu imaginer il y a quelques dizaines d’années.MéthodesExpérimentation en laboratoireL’expérimentation a surtout été utilisée pour étudier les phénomènes de croissance et de compétition entre espèces. La croissance d’une population isolée est généralement bien représentée par une courbe logistique sigmoïde dont les paramètres dépendent à la fois de l’espèce considérée et des facteurs de milieu (fig. 1). L’abondance tend asymptotiquement vers une limite qui est la charge biotique maximale compatible avec le milieu. On a constaté que ces modèles logistiques s’ajustaient bien à la croissance de nombreux organismes, des ciliés aux vertébrés en passant par les insectes et les crustacés.La compétition a été étudiée entre deux espèces différentes occupant la même niche écologique et entre deux mutants de la même espèce. Dans le premier cas, des élevages de drosophiles sur milieu artificiel avec une quantité de nourriture constante ont montré que, suivant les conditions de température et d’humidité relative, l’une des espèces éliminait l’autre ou les deux cohabitaient, l’équilibre final étant indépendant des conditions initiales. Les résultats sont identiques lorsqu’on met en compétition deux mutants. La dynamique et la génétique des populations sont donc étroitement liées.Malgré leur intérêt théorique, la portée de ces expériences reste cependant limitée. Elles sont en effet menées en milieux maintenus artificiellement stables avec une ou deux populations monospécifiques isolées, conditions très différentes de celles que l’on rencontre dans les peuplements naturels dont la complexité ne peut être reconstituée au laboratoire.Échantillonnage des populations naturellesPour l’étude quantitative des populations animales, on ne dispose pas de registres d’état civil ou de recensements détaillés, comme en démographie humaine. Par conséquent, la seule méthode valable pour déceler le sens et l’amplitude d’éventuelles variations consiste à comparer les données d’échantillonnages échelonnés dans le temps ou l’espace. Les résultats seront d’autant meilleurs que les échantillons prélevés seront plus nombreux et représentatifs de la population étudiée. Un échantillon est dit représentatif ou non biaisé lorsqu’il donne une image réduite, mais non déformée, de la population dont il a été extrait. Malheureusement, la plupart des techniques de prélèvement ou de capture sont sélectives vis-à-vis de certaines catégories d’individus: si, par exemple, les jeunes se laissent prendre plus facilement que les adultes, le nombre des premiers sera surestimé par rapport à celui des seconds.Par une stratégie d’échantillonnage judicieuse, le dynamicien doit s’efforcer d’éviter ces biais ou de les réduire le plus possible. Il faut cependant noter que l’utilisation d’échantillons systématiquement biaisés est encore légitime et conduit à des conclusions valables pour autant que le biais reste constant. Ainsi, lorsqu’il s’agit d’une population exploitée, telle qu’un stock de poissons ou de crustacés, la fraction capturée, ou phase exploitée, peut différer largement de l’ensemble de la population existant sur les lieux de pêche, ou phase disponible. Les différences entre la structure démographique de ces deux phases résultent de la sélectivité plus ou moins grande des engins de capture. Il existe en principe des statistiques de prises ou de débarquements tenues à jour et il est facile d’obtenir, à l’aide d’un bon échantillonnage stratifié, par port de débarquement ou par bateau, une représentation très exacte de la phase exploitée et d’en faire une étude dynamique sans connaître les caractéristiques de la population tout entière. Il est dans ce cas préférable de parler de dynamique des stocks exploités plutôt que de dynamique des populations.Modélisation et simulationLa complexité des phénomènes à étudier en dynamique des populations nécessite le recours à des modèles simplifiés, dépendant d’un nombre aussi restreint que possible de paramètres auxquels on donne des valeurs choisies pour que le modèle soit ajusté aux observations. Des solutions analogiques comme l’appareil à billes construit pour représenter le stock de plies de la mer du Nord en 1958 à Lowestoft par I. D. Richardson et J. P. Gulland n’ont plus qu’un intérêt anecdotique. Les modèles mathématiques sont beaucoup plus performants et faciles à manier. Parmi les plus simples, on peut citer les spirales de Lotka pour représenter les variations d’abondance de deux populations dont l’une vit aux dépens de l’autre comme dans les systèmes proie-prédateur et hôte-parasite; ces spirales obéissent à l’équation B = 切(f ) correspondant à l’équation différentielle:
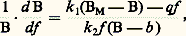 où B est la biomasse, BM la biomasse maximale, f l’effort de pêche, k 1, k 2, q et b des paramètres. Mentionnons encore les modèles de W. B. Schaefer (1954, 1957), fondés sur la proportionnalité inverse entre l’abondance d’un stock et le taux d’exploitation de ce stock, dont on déduit facilement la production équilibrée. Grâce aux ordinateurs, les dynamiciens peuvent utiliser des modèles de plus en plus sophistiqués pour aborder la dynamique des populations auxquelles les modèles simples ne s’ajustent pas suffisamment bien.Lorsqu’on fait varier les paramètres dont ils dépendent, les modèles permettent de simuler le devenir d’une population dans une infinité de situations fictives. Parmi celles-ci, il en est qui se sont déjà réalisées et d’autres qui ont des chances de se réaliser dans les années à venir. La simulation permet donc de tester l’adéquation des modèles à décrire des états déjà observés aussi bien qu’à prévoir des états futurs lorsque la population se trouvera soumise à tel ou tel facteur.Analyse des cohortesOn appelle cohorte l’ensemble des individus de même âge, ayant vécu dans les mêmes conditions et qui viennent périodiquement grossir ou renouveler l’effectif d’une population. Les cohortes sont donc des classes d’âge qui peuvent être annuelles, trimestrielles, mensuelles, etc. Chacune comporte un effectif initial dit nombre de recrues, qui diminue au cours du temps sous les effets de la mortalité naturelle due à la prédation, aux maladies, à la sénilité et éventuellement de la mortalité due à l’exploitation par l’homme. Le recrutement peut varier considérablement d’une cohorte à la suivante en fonction de l’importance du stock de géniteurs dont elle provient et surtout des conditions de milieu plus ou moins favorables au moment de la reproduction et du développement des œufs ou des jeunes. Selon la durée de vie moyenne des individus et la fréquence des générations, une population peut se composer d’une seule ou d’un nombre variable de cohortes.L’analyse des cohortes est un ensemble de techniques permettant de décomposer une population animale en ses cohortes constituantes et d’étudier leur devenir en recherchant par simulation la valeur du recrutement et des coefficients de mortalité qu’il est nécessaire d’appliquer à chacune d’elles pour que l’ensemble représente aussi exactement que possible les structures déduites d’une série chronologique d’échantillons. On arrive ainsi à prévoir les effectifs des cohortes déjà recrutées jusqu’à leur extinction et la structure de la partie de population correspondante, le recrutement des cohortes à venir restant inconnu et sujet à fluctuations. L’analyse des cohortes a été appliquée avec succès dans un certain nombre de cas, notamment à la dynamique des stocks de thons exploités en vue de fixer des quotas annuels à ne pas dépasser par la flotte de pêche internationale.ApplicationsLutte biologiqueLa lutte biologique a pour objet de contrôler l’abondance d’une population cible, jugée nuisible ou indésirable, par introduction d’un prédateur, d’un parasite ou d’un compétiteur, sans modification du milieu. Elle ne doit théoriquement entraîner ni pollution ni autre atteinte à l’environnement. Elle n’est cependant possible que si l’on connaît une espèce susceptible d’agir efficacement sur la cible, si l’on peut en introduire des quantités suffisantes pour obtenir l’effet escompté, enfin si l’espèce choisie ne risque pas de s’attaquer aux autres espèces présentes lorsque la cible se sera raréfiée ou aura disparu. C’est ainsi que l’on a essayé de détruire les larves de moustiques, agents transmetteurs du paludisme, en acclimatant un peu partout des poissons larvivores comme les gambusies.À côté de résultats décevants, la lutte biologique a enregistré des succès spectaculaires. Des coccinelles introduites en Californie ont jugulé l’invasion des cochenilles, qui étaient devenues un fléau redoutable. D’autres insectes prédateurs se sont révélés efficaces dans la lutte contre le pou de San José en Europe ou l’aleurode floconneuse des agrumes sur la Côte d’Azur. L’inoculation volontaire de maladies microbiennes (insecticides à base de Bacillus thuringiensis ) ou virales (myxomatose pour détruire les lapins de garenne) relève également de la lutte biologique.On a parfois recours à plusieurs espèces dont les actions se complètent ou se renforcent. Ainsi, la quasi-disparition d’une chenille qui dévastait les arbres de Nouvelle-Écosse a été obtenue par l’introduction d’une mouche carnassière du groupe des tachines et d’un hyménoptère prédateur du groupe des ichneumons: la première est surtout efficace lorsque la cible est abondante, le second lorsqu’elle est devenue plus rare.Une autre méthode consiste à introduire des individus stériles afin de réduire la fécondité de la population indésirable et d’aboutir à sa disparition totale. Les premiers essais d’éradication par stérilisation ont été tentés contre un diptère américain, le screwworm, parasitant le bétail. Des mâles stérilisés aux rayons 塚 ont été lâchés à raison de 400 individus par mile carré dans l’île de Curaçao. L’éradication fut complète au bout de six mois. Une campagne dans le sud-est des États-Unis avec des lâchers de 500 mâles stériles au mile carré aboutit à la disparition rapide de l’espèce en 1958-1959. Des essais du même genre ont été tentés contre la mouche domestique, des glossines, des moustiques et divers insectes nuisibles aux cultures. Certains ont été positifs, d’autres sans résultats appréciables, faute sans doute de connaissances suffisantes sur la dynamique et la génétique des populations à contrôler.Gestion rationnelle des stocks exploitésBeaucoup de populations naturelles, dont l’homme tire nourriture ou produits utiles tels que peaux, fourrures, huiles, etc., sont encore exploitées de façon anarchique et souvent abusive. Il en résulte un déclin de ces populations, qu’il s’agisse de gibiers, de poissons, de crustacés ou de mollusques, phénomène qui tend à se généraliser et devient de plus en plus alarmant. Les troupeaux de bisons d’Amérique ont été décimés en quelques décennies. Il en est de même des baleines franches. La sardine du Pacifique, dont, entre 1935 et 1945, on pêchait en moyenne 600 000 tonnes par an, a pratiquement disparu. La liste des stocks exploités menacés d’extermination s’allonge chaque année, alors qu’augmente sans cesse la demande destinée à satisfaire les besoins les plus légitimes de l’humanité en nourriture et protéines d’origine animale.Or, la dynamique des stocks permet justement de fixer le taux d’exploitation qui, dans des conditions données, correspond à la production optimale. Des modèles mathématiques de rendement global équilibré, comme ceux de M. B. Schaefer (de 1954 à 1957, fig. 2), de W. W. Fox (1970), de J. J. Pella et P. K. Tomlinson (1969), ou de rendement par recrue, comme ceux de R. J. H. Beverton et S. J. Holt (1957), de W. E. Ricker (1958), sont couramment utilisés dans ce but par les biologistes des pêches. À condition que le milieu environnant reste stable, ils en déduisent le maximum de prises pouvant être obtenu chaque année (maximal sustainable yield : M.S.Y.) sans compromettre ni l’équilibre ni le maintien du stock à son niveau optimal de production. À partir de ces données, les services nationaux des pêches, ou de préférence des commissions régionales et internationales, fixent chaque année, en tenant compte de la conjoncture socio-économique, la prise totale autorisée (total allowed catch : T.A.C.) répartie sous forme de quotas entre les ayants droit.En réalité, les commissions ou les services techniques compétents réglementent l’exploitation des stocks dont ils ont pris en charge la gestion soit par fixation de quotas, soit par limitation de l’effort de pêche (nombre de bateaux, de licences, de permis), soit par modification du maillage des filets afin d’éviter la destruction d’individus de trop petite taille ou plus simplement par fixation d’une taille minimale de capture autorisée, soit encore par fermeture de l’exploitation dans certaines zones ou durant certaines périodes. Les techniques de simulation évoquées ci-dessus permettent de prévoir les états successifs du stock en fonction de telle ou telle réglementation. Ainsi, les décideurs prennent-ils leurs responsabilités en toute connaissance de cause.Bien que la dynamique des stocks exploités ait encore des progrès à faire, elle est dès maintenant en mesure de fournir les bases nécessaires et suffisantes pour une gestion saine et rationnelle des stocks. En ce domaine, échecs et difficultés proviennent du peu d’empressement de certains à se plier à une réglementation quelle qu’elle soit, et surtout au manque d’autorités capables de faire appliquer les décisions prises, d’exercer une surveillance efficace et de sanctionner les contrevenants. Cette carence est particulièrement notable pour la haute mer où s’applique un droit international vieilli et inadapté aux réalités actuelles.Protection des espèces menacéesLes espèces exploitées par l’homme pour sa nourriture, son profit ou son plaisir ne sont pas les seules menacées de déclin et d’extinction. Beaucoup d’autres ont déjà disparu ou sont en train de disparaître, simplement parce que le milieu qui leur convenait a été altéré ou détruit. Les défrichements et les déforestations pour implanter des cultures vivrières ou industrielles, l’assèchement des zones marécageuses, l’extension des réseaux routiers et des périmètres urbains, sans compter les pollutions de toutes sortes, ont pour effet de morceler les populations animales et de les refouler dans des territoires de plus en plus exigus. Or, lorsque son effectif est trop réduit, une population devient, malgré des mécanismes régulateurs naturels dépendant de la densité, incapable de faire face à d’éventuels accidents d’ordre climatique ou épizootique. Elle est alors condamnée à disparaître à plus ou moins brève échéance.Beaucoup d’oiseaux et de mammifères ne subsisteraient plus aujourd’hui s’ils n’avaient été intégralement protégés ou étroitement surveillés dans des réserves, parcs nationaux ou sanctuaires, créés précisément pour assurer leur sauvegarde. Ainsi, en République sud-africaine, le parc Mountain Zebra a été créé pour sauver le zèbre de montagne dont il ne restait que quelques dizaines d’individus en 1937. Comme l’aurochs, définitivement éteint au XVIIe siècle, le bison d’Europe était strictement inféodé aux milieux forestiers. Il serait actuellement disparu sans le parc de Bia face="EU Caron" ゥowieza en Pologne, où un troupeau de plus de 200 têtes a été reconstitué.Lorsqu’il s’agit du sauvetage de petites populations, la dynamique de celles-ci doit être surveillée de très près, grâce à des recensements périodiques à l’aide par exemple de marquages individuels. Le nombre de naissances et de décès observés permet de savoir si l’effectif diminue, se maintient ou s’accroît. Il faut pouvoir prendre à temps les mesures nécessaires: soigner les malades, fournir un complément de nourriture en cas de disette, capturer une partie des individus en cas d’accroissement intempestif pour repeupler d’autres secteurs, éliminer les individus blessés et incurables, les mâles excédentaires, etc. Il s’agit, on le voit, d’un mode de gestion tout à fait différent de celui des stocks exploités, puisqu’il vise non à obtenir la production optimale, mais à maintenir l’effectif total en équilibre avec le territoire et la nourriture disponibles.La dynamique des populations s’était fixé à l’origine trois objectifs: décrire, expliquer et prédire. Les modèles mathématiques dont elle dispose actuellement permettent une description satisfaisante de tous les phénomènes observés. La recherche des causes a mis en évidence un grand nombre de mécanismes particuliers et a largement contribué à faire progresser nos connaissances dans ce domaine. Ces causes sont les unes intrinsèques, relevant de la génétique des populations et de leurs stratégies adaptatives, les autres extrinsèques et dues aux conditions de milieu. Quant aux prédictions, elles n’ont donné de résultats tangibles qu’à court terme et dans la mesure où les facteurs de causalité restent constants ou ne fluctuent qu’aléatoirement autour d’une moyenne stable. La dynamique des populations ne peut prévoir ni les effets de l’apparition de mutants plus résistants ou mieux adaptés que la population d’origine, ni ceux d’une catastrophe telle que aménagements bouleversant le milieu, cataclysme, épizootie, variations climatiques exceptionnelles, pollution, exploitation excessive, etc. L’un de ces facteurs imprévisibles ou généralement plusieurs agissant en synergie se retrouvent à l’origine de tous les développements excessifs et de toutes les disparitions de populations, que l’on souhaiterait prévoir pour les pallier. Tant que l’homme n’aura pas acquis la maîtrise de ces facteurs ni réussi à en contrôler l’action, la dynamique des populations verra ses applications limitées sur le plan des prédictions pratiques.
où B est la biomasse, BM la biomasse maximale, f l’effort de pêche, k 1, k 2, q et b des paramètres. Mentionnons encore les modèles de W. B. Schaefer (1954, 1957), fondés sur la proportionnalité inverse entre l’abondance d’un stock et le taux d’exploitation de ce stock, dont on déduit facilement la production équilibrée. Grâce aux ordinateurs, les dynamiciens peuvent utiliser des modèles de plus en plus sophistiqués pour aborder la dynamique des populations auxquelles les modèles simples ne s’ajustent pas suffisamment bien.Lorsqu’on fait varier les paramètres dont ils dépendent, les modèles permettent de simuler le devenir d’une population dans une infinité de situations fictives. Parmi celles-ci, il en est qui se sont déjà réalisées et d’autres qui ont des chances de se réaliser dans les années à venir. La simulation permet donc de tester l’adéquation des modèles à décrire des états déjà observés aussi bien qu’à prévoir des états futurs lorsque la population se trouvera soumise à tel ou tel facteur.Analyse des cohortesOn appelle cohorte l’ensemble des individus de même âge, ayant vécu dans les mêmes conditions et qui viennent périodiquement grossir ou renouveler l’effectif d’une population. Les cohortes sont donc des classes d’âge qui peuvent être annuelles, trimestrielles, mensuelles, etc. Chacune comporte un effectif initial dit nombre de recrues, qui diminue au cours du temps sous les effets de la mortalité naturelle due à la prédation, aux maladies, à la sénilité et éventuellement de la mortalité due à l’exploitation par l’homme. Le recrutement peut varier considérablement d’une cohorte à la suivante en fonction de l’importance du stock de géniteurs dont elle provient et surtout des conditions de milieu plus ou moins favorables au moment de la reproduction et du développement des œufs ou des jeunes. Selon la durée de vie moyenne des individus et la fréquence des générations, une population peut se composer d’une seule ou d’un nombre variable de cohortes.L’analyse des cohortes est un ensemble de techniques permettant de décomposer une population animale en ses cohortes constituantes et d’étudier leur devenir en recherchant par simulation la valeur du recrutement et des coefficients de mortalité qu’il est nécessaire d’appliquer à chacune d’elles pour que l’ensemble représente aussi exactement que possible les structures déduites d’une série chronologique d’échantillons. On arrive ainsi à prévoir les effectifs des cohortes déjà recrutées jusqu’à leur extinction et la structure de la partie de population correspondante, le recrutement des cohortes à venir restant inconnu et sujet à fluctuations. L’analyse des cohortes a été appliquée avec succès dans un certain nombre de cas, notamment à la dynamique des stocks de thons exploités en vue de fixer des quotas annuels à ne pas dépasser par la flotte de pêche internationale.ApplicationsLutte biologiqueLa lutte biologique a pour objet de contrôler l’abondance d’une population cible, jugée nuisible ou indésirable, par introduction d’un prédateur, d’un parasite ou d’un compétiteur, sans modification du milieu. Elle ne doit théoriquement entraîner ni pollution ni autre atteinte à l’environnement. Elle n’est cependant possible que si l’on connaît une espèce susceptible d’agir efficacement sur la cible, si l’on peut en introduire des quantités suffisantes pour obtenir l’effet escompté, enfin si l’espèce choisie ne risque pas de s’attaquer aux autres espèces présentes lorsque la cible se sera raréfiée ou aura disparu. C’est ainsi que l’on a essayé de détruire les larves de moustiques, agents transmetteurs du paludisme, en acclimatant un peu partout des poissons larvivores comme les gambusies.À côté de résultats décevants, la lutte biologique a enregistré des succès spectaculaires. Des coccinelles introduites en Californie ont jugulé l’invasion des cochenilles, qui étaient devenues un fléau redoutable. D’autres insectes prédateurs se sont révélés efficaces dans la lutte contre le pou de San José en Europe ou l’aleurode floconneuse des agrumes sur la Côte d’Azur. L’inoculation volontaire de maladies microbiennes (insecticides à base de Bacillus thuringiensis ) ou virales (myxomatose pour détruire les lapins de garenne) relève également de la lutte biologique.On a parfois recours à plusieurs espèces dont les actions se complètent ou se renforcent. Ainsi, la quasi-disparition d’une chenille qui dévastait les arbres de Nouvelle-Écosse a été obtenue par l’introduction d’une mouche carnassière du groupe des tachines et d’un hyménoptère prédateur du groupe des ichneumons: la première est surtout efficace lorsque la cible est abondante, le second lorsqu’elle est devenue plus rare.Une autre méthode consiste à introduire des individus stériles afin de réduire la fécondité de la population indésirable et d’aboutir à sa disparition totale. Les premiers essais d’éradication par stérilisation ont été tentés contre un diptère américain, le screwworm, parasitant le bétail. Des mâles stérilisés aux rayons 塚 ont été lâchés à raison de 400 individus par mile carré dans l’île de Curaçao. L’éradication fut complète au bout de six mois. Une campagne dans le sud-est des États-Unis avec des lâchers de 500 mâles stériles au mile carré aboutit à la disparition rapide de l’espèce en 1958-1959. Des essais du même genre ont été tentés contre la mouche domestique, des glossines, des moustiques et divers insectes nuisibles aux cultures. Certains ont été positifs, d’autres sans résultats appréciables, faute sans doute de connaissances suffisantes sur la dynamique et la génétique des populations à contrôler.Gestion rationnelle des stocks exploitésBeaucoup de populations naturelles, dont l’homme tire nourriture ou produits utiles tels que peaux, fourrures, huiles, etc., sont encore exploitées de façon anarchique et souvent abusive. Il en résulte un déclin de ces populations, qu’il s’agisse de gibiers, de poissons, de crustacés ou de mollusques, phénomène qui tend à se généraliser et devient de plus en plus alarmant. Les troupeaux de bisons d’Amérique ont été décimés en quelques décennies. Il en est de même des baleines franches. La sardine du Pacifique, dont, entre 1935 et 1945, on pêchait en moyenne 600 000 tonnes par an, a pratiquement disparu. La liste des stocks exploités menacés d’extermination s’allonge chaque année, alors qu’augmente sans cesse la demande destinée à satisfaire les besoins les plus légitimes de l’humanité en nourriture et protéines d’origine animale.Or, la dynamique des stocks permet justement de fixer le taux d’exploitation qui, dans des conditions données, correspond à la production optimale. Des modèles mathématiques de rendement global équilibré, comme ceux de M. B. Schaefer (de 1954 à 1957, fig. 2), de W. W. Fox (1970), de J. J. Pella et P. K. Tomlinson (1969), ou de rendement par recrue, comme ceux de R. J. H. Beverton et S. J. Holt (1957), de W. E. Ricker (1958), sont couramment utilisés dans ce but par les biologistes des pêches. À condition que le milieu environnant reste stable, ils en déduisent le maximum de prises pouvant être obtenu chaque année (maximal sustainable yield : M.S.Y.) sans compromettre ni l’équilibre ni le maintien du stock à son niveau optimal de production. À partir de ces données, les services nationaux des pêches, ou de préférence des commissions régionales et internationales, fixent chaque année, en tenant compte de la conjoncture socio-économique, la prise totale autorisée (total allowed catch : T.A.C.) répartie sous forme de quotas entre les ayants droit.En réalité, les commissions ou les services techniques compétents réglementent l’exploitation des stocks dont ils ont pris en charge la gestion soit par fixation de quotas, soit par limitation de l’effort de pêche (nombre de bateaux, de licences, de permis), soit par modification du maillage des filets afin d’éviter la destruction d’individus de trop petite taille ou plus simplement par fixation d’une taille minimale de capture autorisée, soit encore par fermeture de l’exploitation dans certaines zones ou durant certaines périodes. Les techniques de simulation évoquées ci-dessus permettent de prévoir les états successifs du stock en fonction de telle ou telle réglementation. Ainsi, les décideurs prennent-ils leurs responsabilités en toute connaissance de cause.Bien que la dynamique des stocks exploités ait encore des progrès à faire, elle est dès maintenant en mesure de fournir les bases nécessaires et suffisantes pour une gestion saine et rationnelle des stocks. En ce domaine, échecs et difficultés proviennent du peu d’empressement de certains à se plier à une réglementation quelle qu’elle soit, et surtout au manque d’autorités capables de faire appliquer les décisions prises, d’exercer une surveillance efficace et de sanctionner les contrevenants. Cette carence est particulièrement notable pour la haute mer où s’applique un droit international vieilli et inadapté aux réalités actuelles.Protection des espèces menacéesLes espèces exploitées par l’homme pour sa nourriture, son profit ou son plaisir ne sont pas les seules menacées de déclin et d’extinction. Beaucoup d’autres ont déjà disparu ou sont en train de disparaître, simplement parce que le milieu qui leur convenait a été altéré ou détruit. Les défrichements et les déforestations pour implanter des cultures vivrières ou industrielles, l’assèchement des zones marécageuses, l’extension des réseaux routiers et des périmètres urbains, sans compter les pollutions de toutes sortes, ont pour effet de morceler les populations animales et de les refouler dans des territoires de plus en plus exigus. Or, lorsque son effectif est trop réduit, une population devient, malgré des mécanismes régulateurs naturels dépendant de la densité, incapable de faire face à d’éventuels accidents d’ordre climatique ou épizootique. Elle est alors condamnée à disparaître à plus ou moins brève échéance.Beaucoup d’oiseaux et de mammifères ne subsisteraient plus aujourd’hui s’ils n’avaient été intégralement protégés ou étroitement surveillés dans des réserves, parcs nationaux ou sanctuaires, créés précisément pour assurer leur sauvegarde. Ainsi, en République sud-africaine, le parc Mountain Zebra a été créé pour sauver le zèbre de montagne dont il ne restait que quelques dizaines d’individus en 1937. Comme l’aurochs, définitivement éteint au XVIIe siècle, le bison d’Europe était strictement inféodé aux milieux forestiers. Il serait actuellement disparu sans le parc de Bia face="EU Caron" ゥowieza en Pologne, où un troupeau de plus de 200 têtes a été reconstitué.Lorsqu’il s’agit du sauvetage de petites populations, la dynamique de celles-ci doit être surveillée de très près, grâce à des recensements périodiques à l’aide par exemple de marquages individuels. Le nombre de naissances et de décès observés permet de savoir si l’effectif diminue, se maintient ou s’accroît. Il faut pouvoir prendre à temps les mesures nécessaires: soigner les malades, fournir un complément de nourriture en cas de disette, capturer une partie des individus en cas d’accroissement intempestif pour repeupler d’autres secteurs, éliminer les individus blessés et incurables, les mâles excédentaires, etc. Il s’agit, on le voit, d’un mode de gestion tout à fait différent de celui des stocks exploités, puisqu’il vise non à obtenir la production optimale, mais à maintenir l’effectif total en équilibre avec le territoire et la nourriture disponibles.La dynamique des populations s’était fixé à l’origine trois objectifs: décrire, expliquer et prédire. Les modèles mathématiques dont elle dispose actuellement permettent une description satisfaisante de tous les phénomènes observés. La recherche des causes a mis en évidence un grand nombre de mécanismes particuliers et a largement contribué à faire progresser nos connaissances dans ce domaine. Ces causes sont les unes intrinsèques, relevant de la génétique des populations et de leurs stratégies adaptatives, les autres extrinsèques et dues aux conditions de milieu. Quant aux prédictions, elles n’ont donné de résultats tangibles qu’à court terme et dans la mesure où les facteurs de causalité restent constants ou ne fluctuent qu’aléatoirement autour d’une moyenne stable. La dynamique des populations ne peut prévoir ni les effets de l’apparition de mutants plus résistants ou mieux adaptés que la population d’origine, ni ceux d’une catastrophe telle que aménagements bouleversant le milieu, cataclysme, épizootie, variations climatiques exceptionnelles, pollution, exploitation excessive, etc. L’un de ces facteurs imprévisibles ou généralement plusieurs agissant en synergie se retrouvent à l’origine de tous les développements excessifs et de toutes les disparitions de populations, que l’on souhaiterait prévoir pour les pallier. Tant que l’homme n’aura pas acquis la maîtrise de ces facteurs ni réussi à en contrôler l’action, la dynamique des populations verra ses applications limitées sur le plan des prédictions pratiques.
Encyclopédie Universelle. 2012.